Polyethylene (PE), the most common petroleum-based plastic, is expressed as “[CH2–CH2]n” and is widely used in everyday life, with an annual global production of approximately 140 million tons.(1, 2) However, the obvious contrast between the remarkable durability and the short service time of PE products leads to the accumulation of PE waste in the environment, which has generated international concern.(3-6)
Several features of PE make it resistant to biodegradation. Among these features are a) PE’s highly stable C–C and C–H covalent bonds; b) its high molecular weight, which makes it too large to penetrate the cell walls of microbes; c) its lack of readily oxidizable and/or hydrolyzable groups; and d) its highly hydrophobic nature.(7) Since the early 1970s, tests on the biodegradation of virgin PE (unpretreated and without any additives) have been performed under natural environmental conditions, including soils, seawater, sludge, and compost, which harbor a multitude of diverse microbial communities.(8-14) These studies have concluded that the biodegradation of virgin PE is extremely limited in selected mixed microbial communities.(2-7)
It is believed that the environmental degradation of PE can be achieved by photo- or thermo-oxidation and then through the biological activity of microorganisms.(15, 16) When PE samples are pretreated with UV light or thermo-oxidation, depolymerization of the PE chains occurs and results in the formation of low molecular weight products, including alkanes, alkenes, ketones, aldehydes, various alcohols, and fatty acids, which are then further degraded by select microorganisms.(15-18) The pretreatments either decrease the hydrophobicity of the polymer, making it more compatible with microorganisms, or introduce groups such as C═O or −OH, which are more prone to microbial degradation.(19)
In contrast, almost no biodegradation of PE through the biological activities of select microorganisms can be observed without pretreatments.(15) The poor biodegradation of virgin PE has been attributed to the lack of specific plastic-degrading microorganisms with long carbon chain oxidization and depolymerization capabilities. Certain researchers have assumed that the time for microbial evolution, which is required for plastic degradation, might be much longer than the history of PE applications.(20)
To date, researchers are still attempting to discover PE-degrading microorganisms through tests involving plastic-waste-contaminated soils, landfills, compost, and marine life and have isolated several bacterial strains (SI Table S1).(20-39) Some of these strains have shown a moderate ability to use virgin PE as a carbon source based on the characterization of biofilm formation on PE films, weight loss of PE materials, surface deterioration, and changes in the mechanical and thermal properties of PE (SI Table S1). For example, it has been reported that a Pseudomonas strain causes weight losses of up to 20% in the tested PE within 120 days.(32)
Additionally, a Canadian Web site has reported the isolation of two novel bacteria that rapidly degrade 22% or more of the tested PE weight over a six week period.(39) However, these promising reports of PE degradation based on weight loss have not been confirmed due to a failure to provide additional supporting evidence, such as changes in surface hydrophobicity, the formation of oxidized carbonyl groups, the scission of long-chain molecules, or the release of low molecular weight products. No report on the deposits of the aforementioned bacteria in any culture collection center has been provided. More studies are needed to confirm PE degradation by microorganisms via sequential two-step reactions, which have been achieved by the synergistic actions of physicochemical oxidation and through the biological activity of microorganisms, as proposed by Albertsson et al.(15)
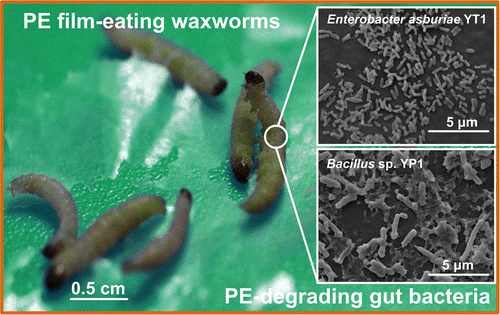
We found that the grain pest larvae of Plodia interpunctella (Hübner), commonly known as waxworms or Indian mealmoths, can damage PE packing films by chewing and eating the film (SI, Figure S1). A similar observation stating that various species of stored-product insect pests like to chew and penetrate PE packaging films has been previously reported.(40) However, the focus of the previous report was to test for and discover a package film material that was resistant to the insects’ destructive tendencies. We were inspired by the insect used in that test and began to wonder whether a microbial symbiont capable of digesting PE was present in the waxworm gut and whether the gut bacteria could act as a source for the isolation of PE-degrading microorganisms.
In this study, with PE film as a sole carbon source, we isolated two PE-degrading bacterial strains from the enrichment of waxworm gut contents and identified that these bacteria were capable of degrading PE within a limited incubation period based on the characterization of biofilm formation, changes in the PE’s physical properties (tensile strength and surface topography), chemical structure (hydrophobicity and appearance of carbonyl groups), molecular weight (accompanied by the formation of daughter products), and weight loss. Our results confirmed PE biodegradation by the two gut bacteria isolated from waxworms and indicated that the bacteria from these plastic-chewing insect larvae were a promising source of plastic-degrading microorganisms.
Materials and Methods
PE Film and Medium
Linear low-density PE (LDPE) film (DFDA-9020, 22.5 μm thickness) was purchased from SINOPEC Beijing Yanshan Company in China. No catalysts or additives were added to this PE product, according to the manufacturer’s standard (Q/SH3180 014). The chemical composition was characterized by a thermogravimetric analyzer (TGA, 209F1, NETZSCH, Germany) and Fourier transform infrared spectroscopy (FTIR, iN10MX, Nicolet, USA). The molecular weight was analyzed using high-temperature gel permeation chromatography (GPC, PL220, Agilent, USA), and the average molecular weight (Mw) and number-average molecular weight (Mn) were found to be 88,200 and 27,700, respectively. The PE film was cut into 50 mm × 50 mm square sheets for incubation in an agar medium plate and small 3 mm × 3 mm pieces for incubation in a liquid medium. The PE materials were weighed, disinfected in 75% ethanol, and then air-dried in a laminar-flow clean bench prior to use. The liquid carbon-free basal medium (LCFBM) prepared with deionized water contained (per 1,000 mL) 0.7 g of KH2PO4, 0.7 g of K2HPO4, 0.7 g of MgSO4·7H2O, 1.0 g of NH4NO3, 0.005 g of NaCl, 0.002 g of FeSO4·7H2O, 0.002 g of ZnSO4·7H2O, and 0.001 g of MnSO4·H2O, according to the ASTM standard for determining the resistance of plastics to bacteria (G22-76, 1996).(41) The carbon-free basal agar medium (CFBAM) was prepared by adding 15 g of agar to 1,000 mL of LCFBM. These media were sterilized by autoclaving at 121 °C for 20 min.
Enrichment and Isolation of PE-Degrading Bacteria
Once the waxworms had chewed enough of the PE bags to cause significant observable damage, approximately 200 of the larvae were collected from the PE bags containing millet grains (SI, Figure S1). To obtain the gut contents for the enrichment of PE-degrading bacteria, the surfaces of the larvae were sterilized via immersion in 75% ethanol for 1 min and then rinsed 2 times with sterile saline water (SW). Next, the guts of the larvae were drawn out and pooled into a 50 mL centrifuge tube containing 40 mL of SW. After being shaken on a vortex mixer for 5 min, the gut tissues were carefully removed with a pipet. The remaining suspension, used as a microbial inoculum, was transferred into a 250 mL Erlenmeyer flask that contained 1 g of the small PE pieces and 80 mL of LCFBM. This flask was incubated on a rotary shaker (120 rpm) at 30 °C.
After 60 days, the residual PE pieces were removed, and the cultures were spread across plates with three different agar media containing complex organic carbon substrates: potato dextrose agar (PDA), bean sprout agar (BSA), and beef extract-peptone agar (BPA). After an incubation period of 24 h, the colonies that formed were transferred to plates with fresh agar medium, where they were kept until pure colonies of isolates (based on a microbial purity test) could be obtained. These isolates have been deposited at the China General Microbiological Culture Collection Center (CGMCCC) in Beijing, China.
Preliminary Screening of Isolates for PE-Degrading Ability
The bacterial isolates were grown in liquid nutrient broth (NB) medium for 12 h. The cells that grew were collected via centrifugation (10,000 rpm) and rinsed with SW to remove the residual medium. This procedure was repeated three times. Next, the collected cells were resuspended in SW to obtain a cell suspension of approximately 108 cells per mL. The melted sterile CFBAM (15 mL) was poured into a Petri dish (90 mm diameter) and cooled to ambient temperature. The cell suspension (0.5 mL) was spread homogeneously across the surface of a CFBAM plate, which was subsequently covered with a PE sheet (50 mm × 50 mm). In the sterile controls, PE sheets were added without inoculation of the cell suspension. CFBAM plates inoculated with the cell suspension without added PE film were used as a control to check whether the isolates could grow on agar alone. All plates were then sealed with Parafilm and incubated at 30 °C and 85% r.h. for 28 days. Three plates were prepared for each isolated strain. The evidence of PE biodegradation was preliminarily determined by characterizing the changes in the tensile strength of the incubated PE sheets on a universal test machine (AGS-X-1KN, Shimadzu, Japan), according to the standard method.(42) To examine changes in the PE film’s hydrophobicity over 28 days, the sole carbon source of the isolated strains was a PE sample in the LCFBM. Afterward, the biofilm on the PE sheet was completely removed by mixing it with 2% w/v aqueous sodium dodecyl sulfate (SDS) for 4 h and then rinsing it with deionized water, according to the washing procedure reported by Sivan A et al.(21, 22) Following the treatment, no cells were observed on the surface of the PE film. The change in the PE surface’s hydrophobicity was determined by measuring the water contact angle (WCA) using a contact angle measuring device (OCA40, Dataphysics, Germany).
Characterization of Biofilm Formation and Cell Viability
The isolates with positive results obtained during preliminary screen tests were further tested for their ability to form biofilms on PE film as a sole carbon source and for the cell viability in the biofilm. A cell suspension (108 cells per mL) was spread across the CFBAM plate, which was subsequently covered with the PE film sheet (50 mm × 50 mm), as described above. Three samples were tested at incubation periods of 0, 3, 7, 14, 21, and 28 days at 30 °C and 85% r.h. For the incubation of zero days, samples were collected after 3 h of inoculation. The plates without inoculation served as a sterile control. After a biofilm formed on the PE surface, the cell number of bacteria that had grown on the PE film surface was counted using the following procedure. The PE film sheet containing a biofilm was transferred into a 50 mL centrifuge tube with 40 mL of SW, and the tube was shaken on a vortex mixer for 5 min. The PE film was transferred into another centrifuge tube with 40 mL of SW, and this tube was also shaken on the vortex mixer for 5 min. The suspensions of the two tubes were then pooled in a centrifuge tube, which was centrifuged at 10,000 rpm for 15 min. The supernatant was discarded, and the pellet was resuspended in 5 mL of SW. The cell number of the suspension was counted using the series dilution method of plate counting. The biofilm on the PE film sheet was examined using a scanning electron microscopy (SEM, Quanta FEG250, FEI, USA),(21) and the viability of the bacterial cells was characterized in situ using a fluorescence microscope (BX51, Olympus, Japan) after staining with the LIVE/DEAD BacLight Bacterial Viability Kit.(22)
Microscale Analysis of Surface Topography and Chemistry
After a 28-day incubation period, the biofilm on the PE sheet was completely removed using a washing procedure with 2% w/v aqueous sodium dodecyl sulfate (SDS) solution for 4 h.(21, 22) The PE sheets in the sterile control were also treated using the same procedure. Consequently, the surface topography of all PE sheets was characterized by SEM observation (Quanta FEG250, FEI, USA) and with an atomic force microscope (AFM, Dimension Icon, Veeco, USA).(21, 26) The surface chemical components were investigated using X-ray photoelectron spectroscopy (XPS, Thermo Scientific, USA) and a microattenuated total reflectance/Fourier transform infrared imaging microscope (Nicolet iN10 MX, Thermo Scientific, USA).(43) During the XPS spectra analysis, scanning was carried out over a broad-band energy range (0–1,200 eV) at an electron takeoff angle of 90° from the sample areas less than 1 mm in diameter. The overlapping peaks in the C 1s region were resolved into their individual components using a peak-fitting program (XPSPEAK v3.0). The view field of the ATR/FTIR microscope was 400 × 400 μm, and the scanning was performed at a step size of 10 μm under a contact pressure of 3 MPa.
Biodegradation Assays
The PE biodegradation was characterized by the timing of its weight loss, molecular weight shift, and the formation of soluble daughter products in the LCFBM. PE pieces (100 mg) and bacterial suspensions (10 mL) were added to a 150 mL Erlenmeyer flask with 40 mL of LCFBM. The final bacterial concentration was approximately 108 cells per mL, and a flask without inoculation served as a sterile control. Three samples were prepared for each incubation period of 7, 14, 21, 28, and 60 days, and the flasks were incubated on a rotary shaker (120 rpm) at 30 °C. For each incubation period, the residual PE pieces were collected, washed using the procedure reported by Sivan et al.,(21, 22) and dried at ambient temperature for the measurement of residual weight. The washed PE residues collected at the end of the test were randomly sampled for molecular weight distribution analysis using the HT-GPC (PL220, Agilent, USA).(23) The liquid culture was centrifuged at 10,000 rpm for 15 min, and the supernatant was filtered through a 0.22-μm membrane filter. The soluble daughter products in the filtrate were analyzed using electrospray ionization-mass spectrometry (ESI-MS, LCQ, Finnigan, USA) in the positive ion mode.(44)
Sequencing and Phylogenetic Analysis
The genomic DNA needed for 16S rDNA amplification of the isolated strains was extracted from cells grown in the late log phase using a conventional proteinase K treatment and phenol-chloroform extraction. Amplification of the 5′ end of the gene was performed with the universal primers 8-F (5′-AGAGTTTGATYMTGGCTCAG-3′) and 1942-R (5′-GGTTACCTTGTTACGACTT-3′). The obtained sequences were aligned with organisms present on the GenBank database using the Basic Local Alignment Search Tool (BLAST) created by the National Center for Biotechnology Information, USA.
Results and Discussion
Preliminary Screen of PE-Degrading Isolates
Eight bacterial strains were isolated during the enrichment of the gut contents of the PE-eating waxworms when using PE as a sole carbon source (SI, Figure S1). Screenings for PE-degrading microorganisms among these isolates were subsequently performed in terms of the colonization on the PE film sheets in the CFBAM plates, the changes in tensile strength of the PE sheets, the floating behavior of the PE pieces in the LCFBM, and the WCA of the PE surfaces. The Gram-positive strain YP1, identified by 16S rDNA sequencing as Bacillus sp., grew as relatively thick, opaque colonies visible to the naked eye on the PE film sheet. The Gram-negative strain YT1, identified as Enterobacter asburiae, grew as visible, thin, translucent colonies (Figure 1a and Table 1). The other six isolated bacterial strains did not develop visible colonies on the PE film sheets. In addition, no visible colonization occurred on the two control plates: a) the CFBAM with a PE film sheet but without inoculation or b) the CFBAM without a PE sheet but with inoculation.
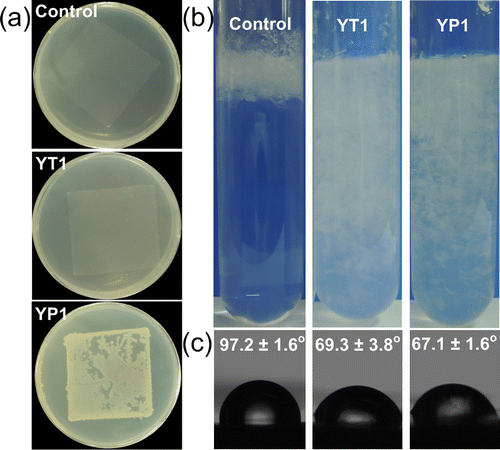
Table 1. Isolation and Identification of PE-Degrading Bacterial Strains

b – BPA = beef extract-peptone agar; PDA = potato dextrose agar.
After a 28-day incubation period, all of the incubated PE film sheets were collected to test their tensile strength. The samples spread with the culture of Bacillus sp. YP1 or E. asburiae YT1 exhibited tensile strength decreases of over 50% compared to that of the sterile controls, while the samples with the other six isolated bacterial strains exhibited no reduction in tensile strength (SI, Figure S2) At the beginning of incubation, the PE pieces floated in the upper portion of the liquid medium because its density (0.915–0.940 g/cm3) was lower than that of water (1 g/cm3) (Figure 1b).
After a 28-day incubation period with strains YT1 and YP1, the PE pieces did not float and became suspended in the LCFBM, whereas the PE pieces in the sterile controls continued to float (Figure 1b). The PE film pieces incubated with the other six strains also continued to float in the liquid medium (data not shown). This result was likely because the cells in strains YT1 and YP1 were able to adhesively grow on the PE species, and this cell adhesion increased the weight of the PE species, resulting in the formation of a suspension.
An analysis of the water contact angles of the PE pieces showed that the WCAs of the samples inoculated with strains YT1 and YP1 were 69.3 ± 3.8° and 67.1 ± 1.6° (n = 5), respectively, which were much lower than the control WCA of 97.2 ± 1.6° (n = 5) (Figure 1c). The other six isolated strains did not significantly change the WCA of the PE pieces (data not shown). The results indicated that the inoculation of bacterial strains YT1 and YP1 decreased the hydrophobicity of the tested PE materials. As the PE materials became less hydrophobic, they became less resistant to subsequent degradation by the bacterial cells.(26) Based on these results, we selected the E. asburiae strain YT1 and Bacillus sp. strain YP1 as potential PE-degraders for further study. Sequences of the two strains were deposited in GenBank under references KJ466896 to KJ466897.
Biofilm Formation Process on PE Film
The biofilm formation on PE film determines its potential biodegradation because a biofilm enables the microorganisms to efficiently utilize the nonsoluble substrate.(21, 22) The biofilm formation processes by the two select strains on the PE film in this study were characterized by cell number counting, morphotypes, and the biofilm’s viability. The biofilm development on the PE film sheet in a CFBAM plate was monitored by counting the number of cells colonizing on the PE film sheet via series dilution during the 28-day incubation.
The results showed that both strains almost immediately adhered to the PE film sheets and built detectable biofilms within the first 3 h of incubation (Figures 2a-c). The cell density of the initial biofilm of strain YT1 was 3.8 × 107 CFU/cm2. The cell density of strain YP1 was 1.4 × 106 CFU/cm2, which was approximately 1 order of magnitude lower than that of strain YT1. The biofilm densities of strains YT1 and YP1 increased from day 3 to day 21, reaching mature biofilms of 16.0 and 9.4 × 107 CFU/cm2, respectively, on day 28 (Figures 2b and c).
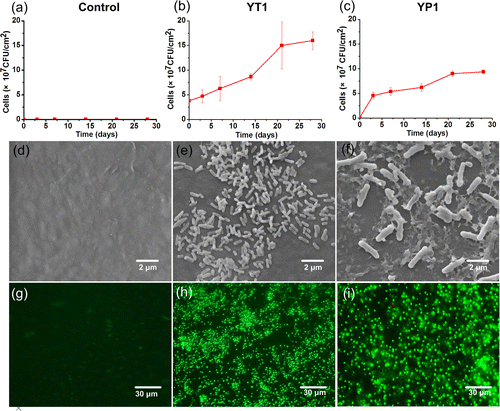
At the end of the 28-day incubation, the biofilm morphotype was observed using SEM. The cells of strain YT1 were rods measuring 0.2 × 0.8 μm. This strain developed a denser biofilm but produced less extracellular polymeric substance (EPS) than the cells of strain YP1 (Figures 2e-f). The cells of strain YP1 were mainly short irregular rods measuring 0.5 × 3 μm, and fewer spore-like cells were observed in its biofilm (Figure 2f). The viability of the biofilms’ bacterial cells grown on the PE film was estimated in situ using a fluorescent microscope after staining with the LIVE/DEAD BacLight Bacterial Viability Kit.(21) Under fluorescence (Figure 2g-i), the live cells (green) dominated in the biofilms of the two strains, and only a limited number of dead cells (red) was observed. The predominance of live cells in the biofilm indicated that these cells received enough growth substrate,(21, 22, 26) which likely came from the metabolism of the PE material and its daughter products.
Changes in Physicochemical Surface Properties
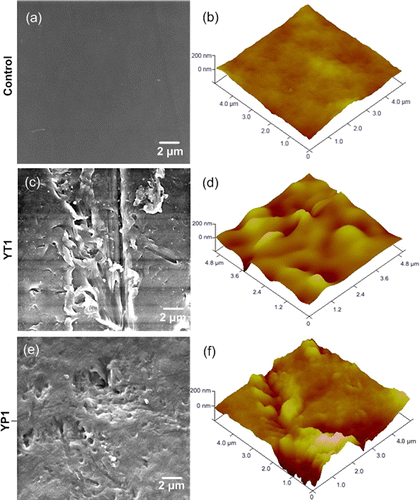
After a 28-day incubation period, the changes in the physicochemical surface properties of the incubated PE film sheets on the CFBAM plates were observed after the biofilm was completely removed from the PE samples. The change in surface topography was examined using SEM and AFM, and the surface deterioration was estimated based on the formation of pits and cavities observed on the surfaces of the PE samples with bacterial inoculation (Figures 3c-f). The surfaces of the controls remained smooth, without any defects (Figures 3a-b). The cavities on the surfaces of the PE sheets had maximum depths of approximately 0.3 and 0.4 μm after inoculation with strains YT1 and YP1, respectively (Figures 3d and f). This type of deep cavity has not been reported by other researchers working on the characterization of PE-degrading microorganisms.(26, 31) The results obtained from the microscale surface topography indicated that the two strains did cause damage to the PE’s physical integrity.
XPS and micro-ATR/FTIR imaging were used to analyze the changes to the surface chemical components and function groups. Figure 4a shows the XPS spectra of C 1s in the inoculated PE sheet surface versus the sterile control. The peaking-fitting result of C 1s for the control showed only one peak at 284.8 eV, which was attributed to a −C–C– group in the long chain skeleton of PE (Figure 4a). In the incubated samples, in addition to the peak at 284.8 eV, another peak appeared at 288.2 eV and was assigned to the carbonyl bands (−C═O−), which is generally considered suggestive of PE oxidation (Figure 4a).(17) Accordingly, the O/C ratios and relative abundance of −C═O– group peaks in the incubated samples are remarkably higher than those of the control (Table 2).
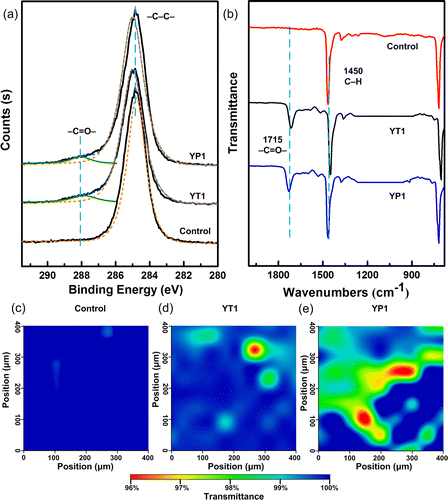
Table 2. O/C Atomic Ratios and Relative C 1s Peak Areas of −C–C– and −C═O– Groups
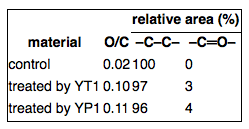
The ATR/FTIR spectra of the inoculated samples’ surfaces were different from that of the control (Figure 4b). All samples had a common peak at 1450 cm–1, which was attributed to the C–H bending vibration in the PE’s long chain skeleton. However, the inoculated samples had a peak at 1715 cm–1, which was assigned to the carbonyl bands (−C═O−). This observation was consistent with the XPS spectra results described above (Figure 4a). Micro-ATR/FTIR images (Figures 4c-e) were constructed based on the transmittance of carbonyl bands (1715 cm–1), with pixels ranging in color from blue to red (transmittance: 100–96%). Compared to the controls, the surfaces of the inoculated samples showed increased amounts of color ranging from light blue to red, indicating heterogeneous oxidation by the bacterial strains. A previous study on PE degradation after pretreatment has demonstrated that the appearance of carbonyl groups is an essential sign of PE biodegradation.(19) Wasserbauer et al. have indicated that PE foils, which were exposed to bacterial (Bacillus brevis and Pseudomonas putida) and liver homogenates, showed oxidation structures and signs of carbonyl-like groups in FTIR spectra.(45) However, these researchers have not reported follow-up work on PE degradation. To the best of our knowledge, no other researchers have reported the appearance of carbonyl groups in the XPS and FTIR spectra of PE foils or films inoculated with microorganisms. In this study, we observed an increasing number of carbonyl group peaks in the samples inoculated with strains YT1 and YP1 using XPS spectra and micro-ATR/FTIR image analysis, indicating that the isolated strains were capable of attacking or oxidizing PE structures to produce carbonyl groups without any pretreatment, such as photo- or thermo-oxidation.
Weight Loss and Molecular Weight Decrease of PE Samples
PE’s degradation efficiency can be directly measured by the weight loss of samples, while the material’s depolymerization can be documented by the shift of its molecular weight and the release of lower molecular weight compounds. The weight loss over time of the PE samples inoculated with the two strains is presented in Figure 5a. During a 60-day incubation period in LCFBM, the weight loss of the PE specimens with inoculation increased consistently, whereas that of the controls did not (Figure 5a). On day 60, the incubation with strain YP1 resulted in a net loss in the PE sample of 10.7 ± 0.2%, which was higher than the 7.5% weight loss reported over a 56-day period for Rhodococcus rubber C208, a previously studied PE-degrading strain.(21-23) Other researchers have reported that incubation with four PE-degrading Bacillus strains results in a net loss in the samples of only up to 5%, based on our estimate of their 60-day removal efficiency.(26, 28) In this study, the incubation with YT1 resulted in a net weight loss of 6.1 ± 0.3%. To date, this is the first reported Enterobacter strain with the ability to degrade PE in the literature. The growth yields of YP1 and YT1 on PE as a sole carbon source were estimated to be 0.82 ± 0.05 and 0.66 ± 0.04 g cells per g PE, respectively.
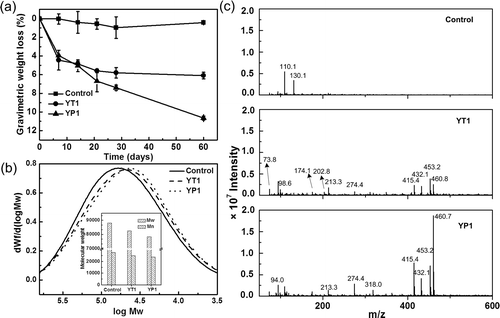
The molecular weight and molecular weight distribution (MWD) of the PE samples after a 60-day incubation were determined using HT-GPC. The MWDs of the PE samples incubated with strains YT1 and YP1 showed a clear negative trend compared to the controls (Figure 5b). The molecular weights (Mw/Mn) of the incubated PE samples containing strains YT1 and YP1 were 82,500/24,700 and 78,200/23,900, respectively, which represents a ∼6–13% reduction from that of the control (Figure 5b internal figure). The decrease in molecular weight suggested that depolymerization/cleavage of the PE’s long chain structure occurred, and lower molecular weight fragments were formed in the presence of these bacterial strains. The formation of water-soluble daughter products was characterized by ESI-MS.(44) The control exhibited a simple spectrum with only two peaks at m/z 110.1 and 130.1. In the samples inoculated with the two strains, 12 new peaks appeared, covering an m/z range from 100 to 600 (Figure 5c).
Most of these compounds contained elements of C, H, and O in their assumed formulas (SI, Table S2). Fold-change analysis was used to identify these 14 compounds with differential abundances in each of the incubated samples versus the products released from the controls (SI, Table S2). In addition to the two compounds at m/z 110.1 and 130.1, the other 12 compounds were more abundant in the PE samples inoculated with strains YT1 and YP1 (SI, Table S2). Among these 12 compounds, four with an m/z range between 400 and 500 were ten times more abundant in all incubated samples than in the control (SI, Table S2). These potential daughter products occurred in the media containing an inoculation but were absent in the control. Although the bacterial cells could secrete a diverse mixture of secondary metabolic products in the medium, causing peaks, the ESI-MS results still provide additional information on the occurrence of PE’s heterotrophic microbial metabolism. In summary, the weight loss and molecular weight decrease of the PE samples and the appearance of daughter products indicate that the bacterial strains isolated from waxworm guts were capable of degrading PE.
Implications
To date, photo- and thermal-based pretreatments have been considered a critical step in the biodegradation of PE, and the studies of PE-degrading microorganisms have focused on plastic-contaminated environments, such as soils, compost, landfills, and marine environments.(2) This study is the first to report the presence of PE-degrading bacteria in the guts of waxworms, which eat PE films. PE degradation was confirmed not only by the bacterial growth on PE films, which acted as sole carbon source and the PE substrate’s weight loss, but also by identification of key reactions similar to those in photo- and thermal-based pretreatments, such as changes in surface topography, decreases in hydrophobicity, the formation of carbonyl groups, decrease in molecular weight, and release of soluble daughter products. The soluble daughter products were not identified in this study, and future study will be needed to identify the daughter and/or intermediate products of PE degradation and the associated metabolic pathways.
Further research should also focus on the ecophysiological relationship between the PE-degrading bacteria and waxworms, which may be mutually beneficial, similar to the microbial symbionts in cow rumens. The discovery of the presence of PE-degrading microorganisms in waxworm guts has provided a promising direction for further study of the microbial degradation of both PE and other plastics, along with new insights into the life cycle of synthetic plastics in nature. In addition, the isolation and characterization of more PE-degrading microorganisms from this source and a better understanding of the enzymatic system involved in PE degradation could be helpful in the development of remediation approaches for plastic wastes, which could eliminate plastic pollution concerns.
Supporting Information
Summary of the updated reported virgin PE-degrading microorganisms (Table S1). Fold-change analysis of the compounds released from the biodegradation of PE (Table S2). Photograph of waxworms eating PE film (Figure S1). Changes in tensile strength of the PE film samples incubated with eight bacterial strains (Figure S2). This material is available free of charge via the Internet at http://pubs.acs.org.
Author Contributions
J.Y. and Y.Y. contributed equally to this work. The authors declare no competing financial interest.
Acknowledgment
This research was supported by the National Natural Science Foundation of China (51373006, 20477002), the State Basic Research Program of China (2014CB931800), and the Shenzhen Key Laboratory of Bioenergy (grant CXB201005240001A). The authors thank Ms. Andria Wu for her help with manuscript preparation.
Source
American Chemical Society (ACS), press release, 2014-11-10.
Supplier
American Chemical Society (ACS)
Beihang University Beijing
Beijing Genomics Institute-Shenzhen (BGI-Shenzhen)
National Natural Science Foundation of China
Stanford University
Share
Renewable Carbon News – Daily Newsletter
Subscribe to our daily email newsletter – the world's leading newsletter on renewable materials and chemicals







